Without water, the body will shut down its need to ingest food
A new study identifies sensors in the gastrointestinal tract that signals the body that food ingestion is no longer required
Bethesda, MD – According to the United Nations, 14 African countries now face water scarcity or water stress. By the year 2025, experts agree that eleven more countries will join the list and half the world will live with water stress.
The consequences of water drought can be terrible – a loss of livestock and crops can lead to overall starvation of a nation’s population. International relief agencies may respond with necessary food supplies. But their largesse in offering food may be for naught, for a new study suggests that without water, the body’s physiology will cause the body to involuntarily reduce feeding, leading to dehydration or anorexia. The findings suggest that deprivation is mediated by a sensor located in either the gastrointestinal tract or in the mesenteric veins draining the gut. In the absence of drinking water, signals from this sensor provoke the early termination of a meal.
The authors of the study, “Reduced Feeding During Water Deprivation Depends on Hydration of the Gut,” are Guus H. M. Schoorlemmer and Mark D. Evered, both in the Department of Physiology, College of Medicine, University of Saskatchewan, Saskatoon, Canada. Their findings are published in the November 2002 edition of the American Journal of Physiology – Regulatory, Integrative and Comparative Physiology, a publication of the American Physiological Society (APS).
Background
Humans join rats, dogs, cows, camels, and other mammals in reducing food intake during water deprivation. This inhibition of eating assists body fluid regulation in two ways. First, there is normally a large volume of osmotically sequestered water in the gut. Inhibition of feeding allows absorption of this water. Second, the reduced solute load or dissolution of food reduces urinary water loss.
There are several ways in which water deprivation might inhibit food intake. First, drinking is normally associated with eating and disrupting that pattern might inhibit food intake. Second, water deprivation causes a dry mouth, making eating more difficult, especially when the period of water deprivation is long and the food is dry. Third, osmoreceptors (a receptor in the central nervous system that receives stimulus from the blood) or other detectors of body fluid status in the brain, circulation, or gut may be involved. In the rat, various stimuli known to cause thirst and antidiuresis also inhibit feeding, including injection of hypertonic solutions into the gastrointestinal tract or peritoneal cavity, injection of hyperoncotic colloid under the skin, and injection of angiotensin in the brain.
The research team designed a series of experiments to investigate how reducing water consumption reduces food intake. The first step was to measure the effect of short periods of water deprivation on food intake and eating pattern. To investigate the role of plasma tonicity in feeding during water deprivation, they compared changes in plasma tonicity caused by ingestion of food in the presence and absence of drinking water. They then investigated the effect of changes in plasma tonicity, induced by intravenous infusion of hypertonic and hypotonic solutions, on feeding in the absence of drinking water. To examine whether drinking fluids is necessary for normal food intake, the team deprived rats of drinking water overnight while slowly infusing the water these rats normally drank into the stomach.
Methodology
In freely feeding Long-Evans rats, meals were separated by intervals of one hour during the dark period and four hours during the light period. To obtain the meal pattern during water deprivation, they measured food intake every 15 minutes by briefly removing the food hopper, weighing it, and returning it to the cage. Rats were anesthetized and given a postoperative subcutaneous injection of the analgesic buprenorphine-hydrochloride. Tubes were inserted in the vena cava, portal vein, gastric cavity, and intestine, all in a single session. Rats were allowed at least ten days to recover. Experiments began only after rats had regained preoperative body weight.
Infusions were done in the rat’s home cage unless specified differently. The polyethylene infusion tubing was connected to the elbow on the rat’s back and led through the top of the cage. Sterile distilled water or 0.3 M NaCl solution was infused with a disposable syringe mounted in a calibrated pump. For long infusions, the tubing was protected with a metal spring and was connected through a low-friction watertight fluid swivel.
Five experiments were then conducted as follows:
Experiment 1: Analysis of feeding during water deprivation.
Food intake was measured in 11 rats for 7.5 hours, starting at the beginning of the dark period, because rats normally do most of their eating during the dark period. Every 15 minutes the food hopper was removed from the cage, weighed, and returned to the cage. Drinking water was not available during the test period in six rats, but the other rats were allowed to drink. Two days later the experiment was repeated, but the treatments were reversed.
Experiment 2: Effect of the presence of drinking water on changes in plasma tonicity and blood volume induced by eating.
Because it is difficult to take blood samples in spontaneously feeding rats without interrupting feeding, especially in the dark, we took blood samples from rats that had been food deprived and allowed the rats to eat a normal-sized meal.
Experiment 3: Effect of rapid intravenous infusion of water or 0.3 M NaCl on food intake.
Eight rats with a cannula in the thoracic vena cava were tested for the sensitivity of feeding to changes in plasma tonicity. To measure sensitivity of feeding to a fall in plasma tonicity, these rats were deprived of drinking water for 18 hours. Then food was removed, cannulas were connected, and sterile water was infused into the vena cava at a rate of 1.1 ml/min. Food (but not drinking water) was returned six minutes after the end of the infusion.
Experiment 4: Effect of slow infusions of water in the stomach of rats not allowed to drink.
To determine whether the act of drinking is necessary for normal food intake, eight rats with gastric cannulas were deprived of water. During this period, water was infused intragastrically.
Experiment 5: Effect of infusion of water in various body locations on food intake of rats not allowed to drink.
Water was infused in various locations throughout the body to locate the sensors that mediate the effect of hydration on feeding.
Results
The researchers found that:
· Water deprivation reduced meal size, including the first meal.
· Intravenous infusions of 0.3 M NaCl that increased plasma tonicity and sodium concentration to levels greater than those seen after eating caused little or no reduction in food intake.
· Feeding in rats not allowed to drink was restored when amounts of water similar to that normally drunk were infused slowly into the stomach, jejunum, or cecum but not when the water was infused into the vena cava or portal vein.
Conclusions
The findings show that sensors responsive to hypertonicity of the gastrointestinal tract can inhibit feeding. The researchers believe that sensors specifically responsible for the reduction in food intake during water deprivation to be a subpopulation of these located in the proximal gut, because this is the region most likely to be affected by the ingestion of normal size drinks. These sensors are probably in the stomach, in the first part of the small intestine, or in the vasculature that drains these areas, before the liver. In the absence of drinking water, signals from these sensors provoke an early termination of the meal.
These findings are consistent with the view that postingestive, preabsorptive, negative-feedback information from the gut has a primary and direct role in the control of feeding. They suggest that hydration of the gastrointestinal tract is one of the important variables monitored by the gut mucosal receptors and that the information is carried by afferent fibers to the caudal brain stem rather than through changes in the tonicity of the circulation monitored by forebrain or other nonsplanchnic sites. Information from splanchnic sensors monitoring gut hydration may also play a role in the control of drinking, release of vasopressin and oxytocin, salt excretion, and gastric emptying.
Throughout the remainder of this century, the world’s population will be plagued by global warming and continuous drought. Ensuring that the population of the affected countries is not subject to starvation will be a top priority for the international community. These findings will contribute to the development of an effective strategy.
Source: November 2002 edition of the American Journal of Physiology – Regulatory, Integrative and Comparative Physiology, a publication of the American Physiological Society (APS).
The American Physiological Society (APS) was founded in 1887 to foster basic and applied science, much of it relating to human health. The Bethesda, MD-based Society has more than 10,000 members and publishes 3,800 articles in its 14 peer-reviewed journals every year.
Media Contact
More Information:
http://www.faseb.org/aps/All latest news from the category: Health and Medicine
This subject area encompasses research and studies in the field of human medicine.
Among the wide-ranging list of topics covered here are anesthesiology, anatomy, surgery, human genetics, hygiene and environmental medicine, internal medicine, neurology, pharmacology, physiology, urology and dental medicine.


Newest articles

Silicon Carbide Innovation Alliance to drive industrial-scale semiconductor work
Known for its ability to withstand extreme environments and high voltages, silicon carbide (SiC) is a semiconducting material made up of silicon and carbon atoms arranged into crystals that is…
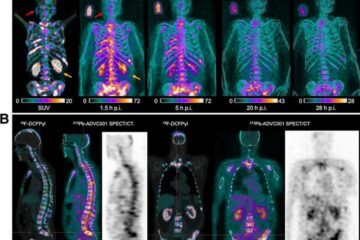
New SPECT/CT technique shows impressive biomarker identification
…offers increased access for prostate cancer patients. A novel SPECT/CT acquisition method can accurately detect radiopharmaceutical biodistribution in a convenient manner for prostate cancer patients, opening the door for more…

How 3D printers can give robots a soft touch
Soft skin coverings and touch sensors have emerged as a promising feature for robots that are both safer and more intuitive for human interaction, but they are expensive and difficult…

