How we sense microbes: Genetic Dissection of innate immunity in insects and mammals
The fundamental challenges.
Infections have always been among the principal causes of death in the human species. In the developed world, microbes may be perceived as “manageable” because sanitation, immunization, and antibiotics hold them in abeyance. But great plagues may lie before us, and our practical success in the prevention and treatment of infection belies the fact that we have much to learn about how immunity operates.
The 20th century witnessed tremendous progress in our understanding of adaptive immunity: the lymphocyte-based system of clonal reactivity and expansion that leads to the production of antibodies by B cells, and to the proliferation of cytotoxic T cells that hold infection in check. Adaptive immunity is remarkable because of its “anticipatory” quality (antibodies can be fashioned against almost any molecular structure including those that do not exist in nature), its exquisite specificity, and because of memory: the ability to mount a faster and stronger response to foreign molecules that we have encountered previously. But immunologists have known that much of our resistance to microbes has nothing at all to do with lymphocytes. Rather, resistance to most microbes is heritable and multifaceted, depending upon neutrophils, macrophages, other phagocytic cells, and upon widely expressed cell-autonomous immune processes. Collectively, this heritable system of defense has been termed innate immunity.
The molecular basis of innate immune recognition remained obscure until recently, and was sought both in insects and in mammals. The small number of molecules that form microbe detection systems, and their clear importance in the initiation of powerful inflammatory responses, have surprised many of us. Moreover, the new understanding as to how infections are perceived by the host within the first minutes following inoculation has wrought major changes in the science of immunology.
Drosophila host defense: a paradigm for innate immunity
Since the beginning of the 20th century, it has been known that insects are highly resistant to microbial infections. Furthermore, it had been established at that time that septic injury leads to the appearance in the cell-free hemolymph (blood) of a significant antimicrobial activity. It took a half century until the first molecules accounting for this inducible antimicrobial activity were characterized in immune-challenged pupae of the Lepidopteran species Hyalophora cecropia by H. Boman on Sweden. These molecules turned out to be cationic, membrane-active peptides. Studies in other insect species and, more generally, on a variety of metazoans including mice and humans, have since shown that the synthesis of antimicrobial peptides is a general phenomenon of the innate host defense.
In the early nineties, the Hoffmann laboratory started the investigation of the antimicrobial defense of Drosophila, with the hope of using the remarkable possibilities offered by Drosophila genetics to decipher the molecular mechanisms of this efficient defense. Relying on a combination of physico-chemical methods and molecular genetics, they first set out to identify the antimicrobial molecules induced in response to septic injury in the fly. They were able to identify seven distinct inducible antimicrobial peptides (or peptide families). These molecules appear to be structurally diverse with activity spectra directed against fungi or against Gram-positive and/or Gram-negative bacteria. The predominant site of biosynthesis of these peptides is the fat body, a functional equivalent of the mammalian liver. Following their synthesis, the peptides are secreted into the circulating hemolymph where their total concentrations can reach values as high as 0,4 mM. It is assumed that the combined activities of the seven distinct groups of antimicrobial peptides largely contribute to the successful blocking of the growth of the invading microorganisms.
When Hoffmann and colleagues analysed in the early nineties the promoter regions of the gene encoding the antimicrobial peptide diptericin, they noted the presence of nucleotide motifs similar to mammalian binding sites for the inducible transactivator NF-kB, reportedly one of the major regulators of immune gene expression in mammals. Mutation of the Drosophila nucleotide sequences similar to NF-kB response elements abolished the inducibility of the diptericin gene. Subsequent studies established that the presence of functional NF-kB response elements is a general phenomenon for the antibacterial peptide genes and in several instances their mandatory roles for challenge induced expression of these genes were confirmed.
These findings raised the tantalizing question whether an equivalent of an NF-kB based transcriptional control is involved in the antimicrobial response of Drosophila. At that time, it had been established that Drosophila produces a member of the NF-kB family, namely the protein Dorsal, which is involved in dorsoventral patterning in the early embryo. Dorsal was also known to be retained in the cytoplasm by binding to the NF-kB inhibitor Cactus. Furthermore, dissociation of Dorsal from its inhibitor, allowing for nuclear translocation of Dorsal and subsequent gene regulation, was understood to be dependent on activation of the transmembrane receptor Toll by a proteolytically cleaved form of the cysteine-knot cytokine-like protein Spaetzle.
The striking similarities between the activation of Dorsal during dorsoventral patterning in the Drosophila embryo and the cytokine-induced activation of NF-kB during inflammation in mammals (e.g. IL-1) induced the group to undertake an in-depth analysis of the antimicrobial defenses in flies mutant for genes of the embryonic regulatory cascade. A pivotal outcome of these studies, published in 1996, was the demonstration that the gene regulatory cascade Spaetzle-Toll-Cactus controls the resistance to fungal infections. It was later understood that this pertains also to the resistance to Gram-positive bacterial infections. This result had an exciting implication. The Toll receptor was indeed known to combine an extracellular domain of leucine-rich repeats to an intracellular Toll-Interleukin-Receptor (TIR) homology domain. LPS had been shown to bind to a leucine-rich repeat domain in the membrane protein CD14, and the TIR domain was known to signal to NF-kB in the case of IL-1. CD14 being glycosylphosphoinositide-anchored, cannot signal by itself to NF-kB. By demonstrating that the combination of an extracellular leucine-rich repeat domain to an intracellular TIR domain can mediate an NF-kB dependent immune response, the Drosophila data provided a conceptual framework for the control of innate immune defenses dependent on genome-encoded (versus rearranged) receptors of microbial ligands.
It had meantime become apparent that mammals produce a family of Toll-like receptors and a new field of study had opened in immunology. Although their existence was reported as early as 1994, such receptors had no known function in mammals. One year after the publication of the role of Toll in the antifungal response of Drosophila, C. Janeway and R. Medzhitov, with whom our group collaborated in the framework of a Human Frontiers in Science Program, reported the existence of a human homologue of Toll and showed that the intracytoplasmic domain of this Toll-like receptor could signal to NF-kB when transfected into a cultured cell line. Significantly though, Janeway and Medzhitov had transfected a contruct in which the extracytoplasmic domain was a CD4 ectodomain consisting of immunoglobulin folds, and their studies did not reveal the receptor's specificity or function.
In parallel to the development of the studies of mammalian TLRs, the Hoffmann group pursued their interests in Drosophila Tolls which by then had appeared to number nine distinct family members. They were initially excited about to the idea that these individual Tolls could bind distinct microbial ligands and signal to NF-kB to direct the expression of various immune-response genes. However, it became clear that -at least as far the control of antimicrobial peptide genes in the systemic response is concerned- only Toll itself plays a role. All Toll receptors, it should be stressed, have tightly regulated, tissue -and stage- specific patterns of expression during embryogenesis and in larval development and are primarily involved in various facets of development. Apparently, only Toll amongst the whole family, has been recruited to actively direct expression of antimicrobial peptides destined to be released into the blood of the fly (systemic immune response, as opposed to epithelial barrier response). Furthermore, the data indicated that Toll activation during the immune defense of Drosophila is mediated by the processed form of the cytokine Spaetzle, but not by direct interaction with microbial ligands. In the terminology coined by the late C. Janeway, Toll does therefore not qualify as a bona fide “pattern recognition receptor”, in contrast to the situation which prevails for mammalian TLRs (see below).
The latter observation raised the question as to which protein served the recognition function of microbes during infection. It was only in 2001 that the Hoffmann group was able to generate a mutant fly which failed to activate Toll in response to Gram-positive bacterial infection. The unbiaised ethyl-methyl-sulfonate induced mutation in this fly line appeared to affect a gene encoding a member of a family of proteins initially discovered in the blood of Lepidoptera through its binding to bacterial peptidoglycan, hence the name of Peptidoglycan Recognition Proteins ( PGRP). The Drosophila genome contains 13 genes coding for members of the PGRP family and the newly generated mutation which affected recognition of Gram-positive bacteria, had changed a conserved cysteine to a tyrosine in the family member PGRP-SA. Interestingly, mammals including humans, also express PGRPs. Mice in which genes encoding PGRP members have been invalidated, show discrete immune phenotypes which are however not of the magnitude with which knock-outs of some of the Drosophila PGRP family members impound on the host defense.
As early as the mid-nineties the studies of the Hoffmann laboratory had further revealed that the defenses to Gram-negative infection in Drosophila rely on a pathway independent from Toll which the authors named Imd (for Immune deficiency) pathway. The end effect of a complex intracellular signaling cascade, initiated by binding of Gram-negative bacterial peptidoglycan to a transmembrane receptor (PGRP-LC), is the phosphorylation and cleavage of the NF-kB family member Relish: this transactivator carries an inhibitory Cactus (IkB)-like domain in its C-terminal region and is evocative of the mammalian NF-kB family member p105. This pathway comprises gene products homologous (or similar) to mammalian RIP, FADD, the MAP3 kinase TAK1, IKKb and IKKg/NEMO. This contrasts with the Drosophila Toll signaling cascade, which lacks all the partners listed above. Overall, the Imd pathway is evocative of the mammalian TNF-µ receptor pathway and strikingly, overexpression of genes encoding upstream members of the Imd pathway can led to apoptosis of immune-responsive cells.
The sum of the data accumulated so far leads to a picture in which two distinct pathways regulate the expression of immune-responsive genes: the Toll pathway, which is triggered primarily by fungal and Gram-positive bacterial infection, and the Imd pathway which serves predominantly in the defense against Gram-negative bacterial aggressions. It should be stressed that each pathway controls the expression of several hundreds of genes. This list includes, but is by no means restricted to, the genes encoding the identified antimicrobial peptides. More than half of the induced genes are of unknown function, underlining that our understanding of the host defense is still very fragmentary.
The prevalent impression which has emerged is that the Drosophila and the mammalian immune pathways have evolved from a reduced number of common ancestral building blocks to their present configurations. Whether the parallels result from convergent evolution or reflect a common ancestry is difficult to decide at present.
Perception of microbes in mammals.
From the mammalian perspective, a different line of investigation was followed, but one with clear parallels to the insect studies described above, and one that converged with the insect research in a remarkable way. Mammals had been viewed as a model for the study of human infectious diseases for more than a century, and in the 1890s, Pfeiffer and Koch began to decipher the inherent toxicity of microbes and molecules derived from them. They identified a heat-stable toxin derived from Gram-negative bacteria, capable of causing shock and death in guinea pigs, mimicking the effects of an authentic infection. Termed “endotoxin,” this substance was ultimately found to be equivalent to lipopolysaccharide (LPS), the major glycolipid constituent of the outer leaflet of the outer membrane of Gram-negative bacteria. Herein lay the seeds of much future discovery, because a microbial molecule of defined structure could clearly trigger powerful inflammatory responses through receptor(s) yet unknown.
LPS is neither toxic to insects, nor to most vertebrates, but is toxic to most mammals, including mice and humans. The effects of LPS are mediated by macrophages, and specifically, by macrophage-derived cytokines, which collectively orchestrate the inflammatory response. In 1985, Beutler and colleagues isolated an LPS-induced cytokine now known as tumor necrosis factor, or TNF, and demonstrated its contribution to LPS-induced shock in mice. Subsequently TNF gene regulation was shown to depend upon NF-?B responsive motifs in the TNF promoter region. The TNF mRNA was also responsive to LPS induction, and translational activation of nearly 200-fold was reported, mediated by the 3'-untranslated region of the TNF mRNA and in particular, by the UA rich element common to the TNF mRNA and other mRNA molecules encoding inflammatory cytokines.
These observations provided molecular endpoints to be used in finding the LPS receptor, ultimately responsible for activating the macrophage. In 1990, Richard Ulevitch and his colleagues established that CD14, a glycosylphosphoinositide-anchored cell membrane protein expressed predominantly by macrophages, was important for LPS signaling as well. Because CD14 has no cytoplasmic domain, it was not immediately clear how the LPS signal might be transduced across the cell membrane. Ulevitch and colleagues noted that CD14 was a molecule composed of leucine-rich repeats, commenting on its similarity Toll, among other molecules. The significance of this observation was unclear at the time, however, and played no part in the search for a transmembrane element within an LPS receptor complex that could carry the LPS signal into the cytoplasm. Conventional biochemical methods were pursued in several laboratories in the search for such a molecule, but were unproductive.
The genetic option.
A non-redundant pathway for LPS recognition was shown to exist in the 1960s and 1970s when substrains of LPS-unresponsive mice (C3H/HeJ mice and C57BL/10ScCr mice) were identified, and the resistance phenotype was in both cases ascribed to spontaneous mutations affecting a single locus, eventually known as the Lps locus. All LPS responses were clearly dependent upon a single, crucial molecule. This molecule was widely envisioned as the “LPS receptor,” though over the years, no evidence of such a receptor could be gleaned from binding studies, likely because of its low abundance and the hydrophobic character of the ligand molecule.
Importantly, experiments with the C3H/HeJ mouse revealed that LPS responses were beneficial in the context of a real infection. Mice that could not sense LPS were quickly overwhelmed when injected with small numbers of Gram-negative bacteria (which produce LPS), while mice that could sense LPS readily contained the infection. Both LPS-sensitive and LPS-resistance mice were competent to cope with Gram-positive microbes (which don't produce LPS). Hence, sensing LPS and responding to it during the first minutes following the inoculation of Gram-negative microbes determined the ultimate outcome of infection.
TNF production, measured by bioassay, was taken as the endpoint of the mammalian response to LPS, and used to positionally clone the Lps locus in the Beutler lab. On 2093 meioses, the locus was confined to a genomic interval approximately 2.6 Mb in length. A BAC and YAC contig was assembled to span the critical region. By shotgun sequencing, Beutler and colleagues identified a gene encoding one of several mammalian homologues of Toll: Toll-like receptor 4 (or TLR4). The gene in question was homologous, in part, to the IL-1 and IL-18 receptors. Further sequence analysis revealed that in C3H/HeJ mice, the gene was modified by a point mutation that altered the cytoplasmic domain of the protein. In C57BL/10ScCr mice, the gene was deleted. Hence, TLR4 was found to be essential for LPS sensing.
Like CD14, TLR4 was a leucine-rich repeat protein. The common motif structure suggested the possibility of a cooperative interaction, with both subunits contributing to a common receptor. Moreover, particularly given the immunological function of Toll in Drosophila, discovered in the Hoffmann lab, the identification of TLR4 as the signaling element of the LPS receptor suggested the likelihood that each of the divergent mammalian TLR paralogues might recognize a distinct subset of microbial ligands, activating a common set of molecular events within the cell, and driving a relatively stereotypic innate immune response. A total of five TLR paralogues were known in mammals in 1998, numbered TLR1 though TLR5. Today we know of 10 human TLRs, 12 mouse TLRs, and 13 TLRs in both species combined. And to a remarkable degree, the hypothesis just stated has been supported by experimental observations.
The subsequent knockout of the gene encoding TLR2 revealed a function in sensing bacterial lipopeptides and lipoteichoic acid. In turn, activating ligands for TLR9, TLR3, TLR7 and TLR8, and TLR5 were identified. TLRs 1 and 6 were shown to contribute to TLR2-mediated sensing by forming heterodimeric complexes with TLR2. TLR1 was required for tri-acyl lipopeptide sensing, while TLR6 was required for diacyl lipopeptide sensing. TLR9 detects DNA of either microbial or host origin, TLR3 detects poly I:C (a mimetic of dsRNA), and TLR7 and TLR8 detect ssRNA or nucleotide-based drugs such as resiquimod (only TLR7, and not TLR8, is active in the mouse).
Accessory subunits contribute to the receptor complexes in some cases. For example, TLR4 signaling depends upon MD-2 (a small protein with a hydrophobic pocket for ligand binding), which binds to the TLR4 ectodomain and may be required not only for initial sensing of LPS but also to permit surface expression of TLR4. CD14 assists in TLR2 signaling and is entirely required for perception of smooth (highly glycosylated) LPS via TLR4. CD36 assists in diacyl lipopeptide sensing. Other cofactors for signaling may also exist.
TLRs are believed to operate as functional dimers, and the overall subunit structure of TLRs is believed to be that of a curved solenoid, or horseshoe, and to the present time, crystallographic models of the ectodomains of TLR2/1, TLR2/6, TLR4/MD-2, and TLR3 have been published. In the case of the TLR2 and TLR4 complexes, a rational explanation for ligand engagement has been offered. The exact molecular events that transpire on the cytoplasmic side of the membrane following activation have not been elucidated.
The range of microbes detected by TLRs
While TLRs 1, 2, 4, and 6 were initially found to recognize components of bacteria and fungi, it later became apparent that the TLR4 complex could recognize viral proteins as well (for example, the env glycoprotein of MMTV, the F protein of RSV, and the G protein of VSV). Later it became clear that TLR signaling leads to the detection of viral nucleic acids as well. Particularly in the case of herpesviruses (MCMV and HSV), immunocompromise results from inadequacy of TLR signaling.
The TLRs are therefore responsible for restricting the growth of many different kinds of microbes. Mice lacking all TLR signaling as a result of mutations in the adapter proteins MyD88 and TRIF (see below) often die of opportunistic infections before weaning, and are difficult to maintain as a stock, although it is possible to do so with effort. If they do manage to survive for several weeks, TLR signaling ablated mice become increasingly resistant to spontaneous infection, consistent with the interpretation that adaptive immune function becomes more and more effective at protecting them independent of innate function.
It is fair to conclude that the mammalian TLRs are a major arm of innate immune perception, though not unique in this role. Some microbes appear to be countered by the NOD/NALP proteins, and others by the RIG-I-like helicases (two distinct classes of cytoplasmic sensors). A full consideration of these pathways is beyond the scope of discussion here, but it may be said that a remarkably small number of protein sensors ignite the most powerful inflammatory responses we know, and prevent small infections from growing out of control prior to the initiation of an adaptive immune response. Where TLR signaling is concerned, the same biochemical pathways that assure survival following the introduction of small numbers of microbes mediate shock and death when infection is out of control.
The role of TLRs in adaptive immunity.
Although numerous papers attest to a requirement for TLRs in the activation of an adaptive immune response, and although there is no doubt that TLRs mediate the adjuvant effect of LPS and a few other molecules of microbial origin that have been known to be adjuvants for many decades, there is no essential need for TLRs in the genesis of an adaptive immune response. In mutant mice with no TLR signaling potential, the production of class-switched immunoglobulins proceeds to the extent that total Ig levels are normal. Antibody responses to defined antigens are normal with all adjuvants tested (including complete Freund's adjuvant, which contains whole mycobacteria emulsified in oil ), and so too is the antibody response to live pathogens; likewise the memory B cell response. Allograft rejection is also unimpaired. On this basis, it may be said that while innate immunity truly depends upon TLR activation, adaptive immunity does not.
Forward Genetics in Mice and the Elucidation of TLR Signaling Pathways
Beginning in the year 2000, a phenotype-driven genetic strategy has been pursued to examine TLR signaling pathways, and to look broadly at molecules required for host defense. The forward screening approach has been carried to considerable depth, and has grown increasingly productive as the speed of positional cloning has accelerated due to technical improvements and its cost has declined.
TLRs were initially shown to signal in part via a cytoplasmic adapter protein termed MyD88 and MAL (also known as Tirap). A mutation produced in the Beutler laboratory, termed Lps2, disclosed a new signaling adapter, now known as TRIF, that works independently of MyD88 and Tirap to carry signals from TLR4 and TLR3. Additional work disclosed a fourth adapter, now known as TRAM, which serves only TLR4. When MyD88 and TRIF mutations were combined, it was found that no TLR signaling could occur, and that mice were severely immunodeficient although they retain adequate adaptive immune responses.
Other mutations revealed that the TLR2/TLR6 complex requires CD36 in order to detect some of its ligands; also that TLRs 3, 7, and 9 (the nucleic acid sensing TLRs) all depend upon a multispanning membrane protein called UNC-93B, which may support trafficking of the TLRs to their proper location within the cell. Still another mutation established that CD14, once known as a component of the TLR4 signaling apparatus, participates in TLR2 complex signaling as well. In all, 18 mutations affecting TLR signal transduction have been created in the Beutler laboratory, and most of them have been positionally cloned. Other mutations, identified in screens for susceptibility to mouse cytomegalovirus (MCMV) have allowed the construction of a “map” of those processes that permit mice to contain t
Media Contact
More Information:
http://www.balzan.orgAll latest news from the category: Life Sciences and Chemistry
Articles and reports from the Life Sciences and chemistry area deal with applied and basic research into modern biology, chemistry and human medicine.
Valuable information can be found on a range of life sciences fields including bacteriology, biochemistry, bionics, bioinformatics, biophysics, biotechnology, genetics, geobotany, human biology, marine biology, microbiology, molecular biology, cellular biology, zoology, bioinorganic chemistry, microchemistry and environmental chemistry.


Newest articles
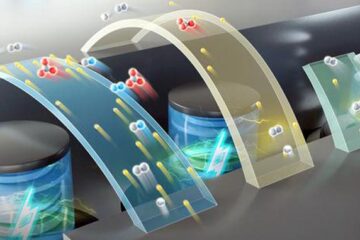
High-energy-density aqueous battery based on halogen multi-electron transfer
Traditional non-aqueous lithium-ion batteries have a high energy density, but their safety is compromised due to the flammable organic electrolytes they utilize. Aqueous batteries use water as the solvent for…
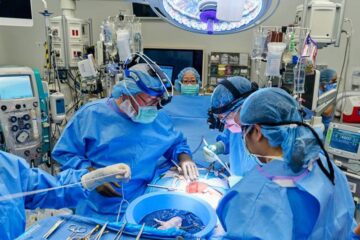
First-ever combined heart pump and pig kidney transplant
…gives new hope to patient with terminal illness. Surgeons at NYU Langone Health performed the first-ever combined mechanical heart pump and gene-edited pig kidney transplant surgery in a 54-year-old woman…
Biophysics: Testing how well biomarkers work
LMU researchers have developed a method to determine how reliably target proteins can be labeled using super-resolution fluorescence microscopy. Modern microscopy techniques make it possible to examine the inner workings…

