Why the hammerhead shark’s head is in the shape it’s in
A comprehensive examination of how the unique head and snout affects maneuverability and the role of its electrosensory function for seeking food along the ocean floor
Why the peculiar head shape of the hammerhead shark developed as it did has been the subject of much speculation. The dorso-ventrally compressed and laterally expanded pre-branchial head is an unmistakable diagnostic feature of the sphyrnid sharks. This unique head shape has been termed the cephalofoil in recognition of its wing-like appearance. The persistence of the sphyrnid cephalofoil over the past 20-25 million years and its presence in several hammerhead shark species of diverse head morphologies tell of its evolutionary success.
Several hypotheses are proposed to explain the evolution of the sphyrnid cephalofoil but few have been empirically tested. Some suggest the cephalofoil acts like a canard to provide hydrodynamic lift and increase maneuvering capabilities. Another hypothesis is that the cephalofoil functions in prey manipulation. Other hypotheses involve potential advantages of spacing sensory structures across the surface or at the lateral ends of the cephalofoil. Another such hypothesis is that pores in the laterally expanded sphyrnid cephalofoil (shark’s head) provide an electrosensory capability that maximizes search area coverage to increase the opportunity to detect food sources.
One of the nation’s leading authors in hammerhead shark physiology has authored three studies that provide insight into these hypotheses. Stephen M. Kajiura, from the Department of Zoology and Hawaii Institute of Marine Biology, University of Hawaii at Manoa, Kaneohe, HI, the researcher behind “Head Morphology and Electrosensory Pore Distribution of Carcharhinid and Sphyrnid Sharks,” originally published in Environmental Biology of Fishes 61: 125–133, 2001; “Maneuvering In Carcharhinid And Sphyrnid Sharks: The Role And Non-Roll of the Hammerhead Shark Cephalofoil” (coauthored by Jesica B. Forni and Adam P. Summers) and Electroreception in Juvenile Scalloped Hammerhead and Sandbar Sharks” (with Kim Holland) Dr. Kajiura will summarize his findings at “The Power of Comparative Physiology: Evolution, Integration and Application” an American Physiological Society (APS) meeting being held August 24-28, 2002, at the Town & Country Hotel, San Diego, CA. For more information about the conference log on to: http://www.the-aps.org/meetings/aps/san_diego/home.htm
Study 1: Head Morphology and Electrosensory Pore Distribution
The enhanced electrosensory hypothesis states that the wider head would sample a greater area, but there would need to be a corresponding increase in the number of electrosensory pores over the wider head area to maintain comparable spatial resolution of small, prey-generated electric fields. Therefore, the enhanced electrosensory hypothesis assumes that sphyrnids have a greater head width than comparably sized carcharhinids and predicts that sphyrnids will have a greater number of pores, which will yield a comparable, or greater pore density. This study compares the distribution of electrosensory pores on two sphyrnids, the scalloped hammerhead, Sphyrna lewini; the bonnethead, S. tiburo; and a representative carcharhinid, the sandbar shark, Carcharhinus plumbeus. Head morphology, pore number and pore density are quantifiable factors to test the assumption and predictions of the enhanced electrosensory hypothesis in hammerhead sharks.
Methodology: Head morphology and the distribution of electrosensory pores were compared between a carcharhinid, Carcharhinus plumbeus, and two sphyrnid sharks, Sphyrna lewini and S. tiburo. Sharks used were incidental mortalities from other research projects and were sampled by gillnet or long line fishing.
Total number of electrosensory pores was counted for 35 S. lewini, 19 S. tiburo, three C. limbatus and 36 C. plumbeus individuals. The size ranges included individuals from juveniles to adults for all species except S. lewini, which included only juveniles. Each head was divided into dorsal and ventral surfaces and all pores were counted on both left and right sides. For the three main study species, S. lewini, S. tiburo and C. plumbeus, four dorsal and eight ventral pore fields were identified based on natural divisions of the pores on the heads.
Results: The greater number of pores distributed on a similar surface area provides S. lewini pups with a higher density of electrosensory pores per unit area compared to C. plumbeus pups. The greater number of ampullae, the higher pore density and the larger sampling area of the head combine to provide hammerhead sharks with a morphologically enhanced electroreceptive capability compared to comparably sized carcharhinids. Both sphyrnid species have a greater head width than the sandbar shark with electrosensory pores distributed across the entire surface of the head for all species. Thus, the electroreceptors are distributed over a greater lateral distance in the sphyrnid sharks.
Conclusions: This study provides evidence that the head morphology of sphyrnid sharks is in line with the assumption and predictions of the enhanced electrosensory hypothesis. The greater number of receptors with an equivalent or higher packing density distributed over a laterally expanded head morphology indicates that the sphyrnid cephalofoil demonstrates the characteristics expected of a head that is optimized for electroreception.
Study 2: Maneuvering in carcharhinid and sphyrnid sharks: the role and non-roll of the hammerhead shark cephalofoil
This study tested whether the anterior foil acts as in canard-winged aircraft to increase maneuverability. This was assessed by hypothesis by determining whether two species of hammerheads (Sphyrna tiburo and S. lewini) turn more sharply, more often, and with greater velocity than a closely related carcharhinid shark (Carcharhinus plumbeus).
Methodology: Video footage of the sharks swimming straight and turning was analyzed to quantify variables of interest. A Hi8 video camera mounted on a sliding track approximately 2m above the surface of the water was used to record the swimming behavior of the scalloped hammerhead and sandbar sharks. The swimming movements of individual sharks were recorded as they swam in a straight trajectory directly under the camera as well as when they made sharp turns (defined as a change in trajectory of > 90°) to orient to a prey-simulating dipole electric field positioned on the substratum directly below the video camera. Video footage of the bonnethead sharks was collected in a similar manner except that a submersible video eye was used as the input to the video camera.
Results: Although the hammerheads were more maneuverable, further investigation revealed that they do not roll their body during turns, negating the possibility that the cephalofoil acts as a steering wing. The findings also demonstrate that hammerhead sharks are more flexible than carcharhinids, and that this flexibility seems due to cross sectional shape rather than number of vertebrae. The two hammerhead species examined exhibited different strategies for high-speed turns: bonnethead sharks use their pectoral fins to steer, whereas scalloped hammerheads use their greater flexibility to power through the turn.
Conclusions: Hammerheads show a greater propensity for executing sharp turns, and maintain a higher speed through the turn. However, the results do not present a complete picture of biologically relevant maneuverability. For example, stopping ability, and carrying velocity through a turn are also mobility related parameters that were not assessed, though they have clear biological relevance. A finer scale study of the flow regimes around the shark’s planning surfaces and has the potential to unravel the specific morphological features that are vital for agile swimming.
Study 3: Electroreception in juvenile scalloped hammerhead and sandbar sharks
This study describes and quantifies the behavioral responses of a sphyrnid and a carcharhinid shark to test the predictions of the enhanced electroreception hypothesis. The responses of juvenile scalloped hammerhead sharks, Sphyrna lewini, and sandbar sharks, Carcharhinus plumbeus, to prey-simulating electric fields were compared to determine if scalloped hammerhead sharks sampled a greater area of the substratum than similarly sized sandbar sharks. The sensitivity of both species to dipole electric fields was also compared. These two species were chosen to represent typical sphyrnid and carcharhinid head morphologies.
Methodology: At the beginning of each trial, a single shark was introduced to the testing arena and allowed to acclimate for several minutes. A well-acclimated shark would swim throughout the entire pen and not limit itself to swimming along the edge of the pen or along the barrier net. To stimulate the shark to start to search for food, an olfactory cue (squid rinse) was introduced to the pen via the odor delivery tube. During each trial, only one of the four dipoles on the acrylic plate was active at any given time while the other three served as controls. When the shark detected the odor and began to demonstrate searching behavior (as indicated by increased tail beat frequency, increased frequency of turning and swimming close to the bottom) the video camera was activated and the shark’s response to the electric field was recorded on videotape at 30 frames per second (fps). A continuous audio commentary of the shark’s movements and behavior was recorded on the audio track of the videotape. After the shark bit at a dipole, that dipole was turned off and another dipole was activated. Trials were brief because the shark would become unresponsive (as indicated by decreased tail beat frequency, decreased frequency of turning and swimming throughout the water column) after a couple of minutes. At the end of each trial the shark was fed to satiation and allowed to rejoin the others on the opposite side of the barrier net.
Results: Thirteen scalloped hammerhead and twelve sandbar sharks were tested for their response to prey-simulating dipole electric fields. When aroused by the food odor stimulus, both species demonstrated a marked change in swimming behavior. The sharks increased their swimming velocity and swam close to the bottom with the ventral surface of the head less than 2cm above the substratum. Both species demonstrated a feeding response by biting at the active dipole and ignoring the non-active dipoles. Sharks sometimes bit repeatedly at an active dipole but immediately stopped biting when the electric current was turned off. Although the hammerhead sharks always responded by biting at the active dipole, the sandbar sharks occasionally did not bite even when they passed within 10cm of the electrodes. These “no bite” responses accounted for 13.3 percent of the total passes. Only responses in which a clear orientation or bite was seen were included in subsequent analyses.
Conclusions: Despite the similarity of response threshold, the orientation pathways and behaviors differed for the two species. Hammerheads typically demonstrated a pivot orientation in which the edge of the cephalofoil closest to the dipole remained stationary while the shark bent its trunk to orient to the center of the dipole. In contrast, sandbar sharks swam in a broader arc toward the center of the dipole. The different orientation patterns are attributed to the hydrodynamic properties of the cephalofoil, which enables the hammerheads to execute sharp turns at high speed. The greater trunk width of the sandbar sharks prevented them from demonstrating the same degree of flexibility. Therefore, although the sphyrnid head morphology does not appear to confer a greater sensitivity to prey-simulating dipole electric fields, it does provide a greater lateral search area, which may increase the probability of prey encounter, and enhanced maneuverability, which may aid in prey capture.
Media Contact
All latest news from the category: Life Sciences and Chemistry
Articles and reports from the Life Sciences and chemistry area deal with applied and basic research into modern biology, chemistry and human medicine.
Valuable information can be found on a range of life sciences fields including bacteriology, biochemistry, bionics, bioinformatics, biophysics, biotechnology, genetics, geobotany, human biology, marine biology, microbiology, molecular biology, cellular biology, zoology, bioinorganic chemistry, microchemistry and environmental chemistry.


Newest articles
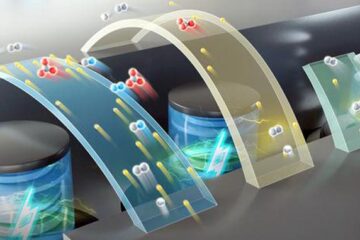
High-energy-density aqueous battery based on halogen multi-electron transfer
Traditional non-aqueous lithium-ion batteries have a high energy density, but their safety is compromised due to the flammable organic electrolytes they utilize. Aqueous batteries use water as the solvent for…
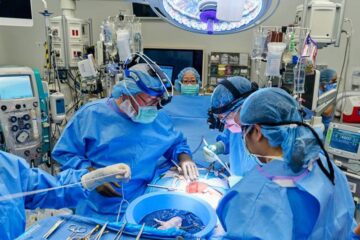
First-ever combined heart pump and pig kidney transplant
…gives new hope to patient with terminal illness. Surgeons at NYU Langone Health performed the first-ever combined mechanical heart pump and gene-edited pig kidney transplant surgery in a 54-year-old woman…
Biophysics: Testing how well biomarkers work
LMU researchers have developed a method to determine how reliably target proteins can be labeled using super-resolution fluorescence microscopy. Modern microscopy techniques make it possible to examine the inner workings…

